Changes in phosphatidylethanolamine composition influence the development of NAFLD and NASH.
About the author
Olga (Olya) Vvedenskaya Sci. Communications Officer
Dr. Dr. Olya Vvedenskaya studied medicine, and further obtained her PhD in the field of molecular oncology. She loves to deliver scientific messages in a clear and accessible manner.
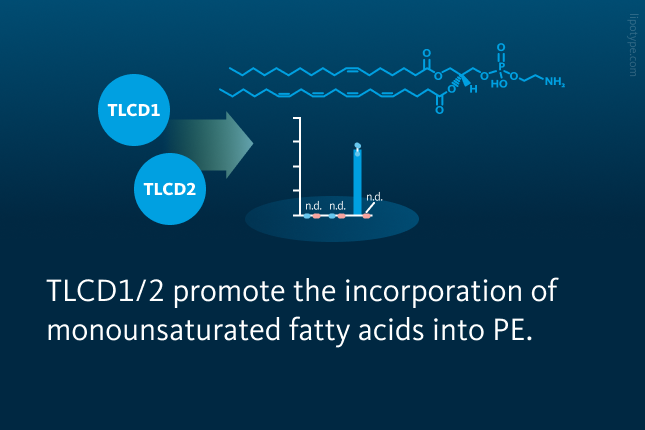
• TLCD1/2 regulate the fatty acyl composition at the sn-1 position of PE • TLCD1/2 promote the incorporation of MUFAs into PE • PE remodelling is linked to the fatty liver disease development
Olga (Olya) Vvedenskaya Sci. Communications Officer
Dr. Dr. Olya Vvedenskaya studied medicine, and further obtained her PhD in the field of molecular oncology. She loves to deliver scientific messages in a clear and accessible manner.
NON-ALCOHOLIC fatty liver disease (NAFLD) is the umbrella term for conditions where lipid excess builds up in the liver, independent of alcohol use. Within NAFLD, there are two main forms. The first is non-alcoholic fatty liver (NAFL), sometimes called “simple steatosis,” where lipids accumulate in the liver, yet there are no signs of inflammation or damage. This stage is often reversible and considered relatively harmless if managed early. The second form is non-alcoholic steatohepatitis (NASH), a more serious condition that involves both lipid buildup and liver inflammation, often leading to scarring (fibrosis) and, in severe cases, cirrhosis or liver cancer.
Recently, experts have updated the terminology to better reflect the metabolic drivers of these conditions. NAFLD is now more and more commonly referred to as metabolic dysfunction–associated steatotic liver disease (MASLD), and NASH is being replaced by metabolic dysfunction–associated steatohepatitis (MASH). For simplicity, in this article, we will continue using the term “fatty liver disease” when referring to NAFLD and its forms (NAFL and NASH), as the authors used these terms in their original publication, while keeping in mind that MASLD and MASH are the more up-to-date medical terms.
While the causes of fatty liver disease are multifactorial, researchers highlight the role of cellular lipid composition, particularly phospholipids in the membranes of liver cells, as one of the key factors in disease progression. In general, the types of fatty acids in membrane phospholipids influence key properties like membrane fluidity, shape, and permeability. These lipids also play a role in how cells respond to oxidative stress and produce signaling molecules. Phosphatidylethanolamine (PE), the second most common phospholipid in mammals, is found mainly in mitochondria and the inner side of the plasma membrane. PE composition impacts metabolism, oxidative stress, and inflammation processes.
Research on PE composition regulation has largely focused on the sn-2 position of PE, where LPLAT enzymes add polyunsaturated fatty acids (PUFAs). These PUFA-rich PEs are prone to peroxidation, which can trigger an iron-dependent form of cell death called ferroptosis. They also serve as precursors for eicosanoids – molecules that are involved in inflammation and other processes. In contrast, the sn-1 position usually contains saturated or monounsaturated fatty acids, but little is known about what controls this or how it affects cell function.
In this study, the researchers used genome-wide association data from mice, loss-of-function models in both mice and human cells, and lipidomic analyses to identify TLCD1 and TLCD2 as key regulators of PE composition. Their findings show that TLCD1 and TLCD2 proteins promote the incorporation of monounsaturated fatty acids (MUFAs) at the sn-1 position of PEs and influence mitochondrial PE composition in mouse liver. Additionally, mice lacking both Tlcd1 and Tlcd2 (double knockout, DKO) displayed reduced progression of liver diseases such as non-alcoholic fatty liver disease and non-alcoholic steatohepatitis when fed a high-fat or Western diet.
To explore genetic control of PE composition, the team analyzed liver PE species in a large genome-lipid association dataset. They identified a strong association between MUFA- and saturated fatty acid (SFA)-containing PEs and a locus on chromosome 11. Within this region, Tlcd1 and Tlcd2 emerged as candidate genes despite being poorly characterized. To test their roles, the researchers generated Tlcd1 knockout, Tlcd2 knockout, and Tlcd1/2 double knockout mice. Lipidomic profiling of liver tissues in controls and mice with fatty liver disease revealed notable changes in PE composition, particularly a significant decrease in MUFA-containing PE species in Tlcd1 knockout mice. The double knockouts showed marked reductions in PEs with palmitoleate (16:1) and oleate (18:1) at the sn-1 position.
Transcriptional control of Tlcd1 and Tlcd2 and their impact on hepatic PE composition n mice. Shown is the complete fatty acyl chain profile of PE species in livers from wild-type and Tlcd1/2 double-knockout (DKO) male mice. The y-axis displays the negative logarithm of p-values from two-tailed multiple t-tests (unadjusted for multiple comparisons). Petkevicius et al., Nat Commun, 13, 6020 (2022), 10.1038/s41467-022-33735-6
To avoid redundancy between Tlcd1 and Tlcd2, and since individual deletions affected PE composition without altering other liver lipids, researchers focused on Tlcd1/2 double knockout mice. In terms of lipidomics changes, these mice showed reduced MUFA and increased SFA content in hepatic PEs, consistent across sex, age, and diet. Despite these changes, DKO mice developed normally and showed no differences in behaviour, energy balance, or glucose tolerance under standard conditions. To explore the mechanism, radiolabeled triolein was used to assess MUFA uptake, which was unchanged in DKO mice, ruling out uptake defects. PE composition was also unaffected by fasting or refeeding, suggesting the changes were not driven by metabolic state.
Primary hepatocytes from DKO mice maintained altered PE profiles in culture, supporting a cell-intrinsic role for Tlcd1/2, meaning that Tlcd1/2 proteins affect PE fatty acid content directly within hepatocytes, and not through changes in diet, circulating lipids, or liver environment. Pulse-labeling showed reduced MUFA and increased SFA incorporation into the sn-1 position of PE, with no changes in PC species. These results indicate Tlcd1/2 specifically promote MUFA incorporation into PE at the sn-1 position through a cell-autonomous, LPLAT-independent mechanism.
Tlcd1/2 facilitate MUFA incorporation into phosphatidylethanolamine in mouse primary hepatocytes. A Experimental design. B Levels of stable isotope–labeled PE and PC species in primary hepatocytes from chow-fed female wild-type and Tlcd1/2 double-knockout mice. Petkevicius et al., Nat Commun, 13, 6020 (2022), 10.1038/s41467-022-33735-6
Most mitochondrial phospholipid metabolism occurs at mitochondria-associated membranes (MAMs) enriched with PE. Microscopy revealed that tagged Tlcd1 and Tlcd2 localize near mitochondria, likely at MAMs, but they are not found within mitochondria themselves, as confirmed by mitochondrial proteome databases.
Lipid analysis of liver mitochondria from Tlcd1/2 DKO mice showed reduced MUFA-containing PE species and increased SFAs at the sn-1 position, mirroring changes seen in total liver lipidomes. Despite PE being synthesized from phosphatidylserine (PS), levels of mitochondrial and total liver PS species were unchanged between genotypes. These findings suggest that Tlcd1/2 interact with mitochondria at the membrane interface and specifically regulate mitochondrial PE composition in the liver.
Tlcd1/2 shape mitochondrial PE composition. Volcano plot showing negative log₁₀-transformed p-values comparing mitochondrial PE species (mol% of total PE) between control and Tlcd1/2 double-knockout male mice (from unpaired two-tailed Student’s t-tests. Petkevicius et al., Nat Commun, 13, 6020 (2022), 10.1038/s41467-022-33735-6
To assess whether changes in mitochondrial PE from Tlcd1/2 deletion affect fatty liver disease, researchers first modelled fatty liver disease-related inflammation in vitro. Primary hepatocytes from the mouse model were treated with lipopolysaccharide and palmitate to induce lipotoxic stress and cytokine production. Tlcd1/2 DKO hepatocytes showed no significant differences in cytokine secretion, de novo lipogenesis, or hepatocyte marker gene expression compared to controls, suggesting no impact on inflammatory response or core cell identity in vitro.
To study the effects of Tlcd1/2 in fatty liver disease development in vivo, male and female Tlcd1/2 DKO mice were fed a high-fat diet (HFD), which induces hepatic steatosis and insulin resistance that does not progress to steatohepatitis and fibrosis. Body weight, body composition, glucose, lipid levels, and fasting insulin were similar across genotypes. However, DKO mice had reduced liver weight, lower hepatic lipid accumulation, and decreased serum alanine aminotransferase (ALT) levels, indicating reduced liver damage. These results suggest that Tlcd1/2 deletion limits HFD-induced NAFLD development, despite no overt systemic metabolic changes.
Tlcd1/2 deficiency lowers liver lipid accumulation and serum ALT activity in HFD-fed mice. Liver weight (relative to body weight), total liver lipid content, and serum ALT activity in high-fat-diet–fed control and Tlcd1/2 double-knockout male and female mice. Petkevicius et al., Nat Commun, 13, 6020 (2022), 10.1038/s41467-022-33735-6
To test if Tlcd1/2 deletion protects against NASH, mice were fed a Western diet (WD). Despite similar weight and glucose tolerance, DKO mice had smaller livers, less hepatic fat, lower ALT levels, and reduced inflammation and fibrosis compared to controls. These effects were absent in chow-fed mice.
DKO mice also showed reduced steatosis and downregulation of genes related to inflammation, immune cell infiltration, and fibrosis. No changes were observed in lipid metabolism genes. Female DKO mice had lower systemic inflammation, with decreased pro-inflammatory cytokines and IL-6, and spleen size. While total liver eicosanoid levels were unchanged, precursor PUFA-containing PE and PC species were altered in DKO livers. These results suggest Tlcd1/2 deletion mitigates NASH progression by reducing fibrosis in males and inflammation in females, possibly due to sex-specific differences in PE metabolism regulated by estrogen-sensitive phosphatidylethanolamine N-methyltransferase activity.
Tlcd1/2 deficiency reduces NASH severity in Western diet-fed mice. A Liver weight (relative to body weight), B Serum ALT activity, C Hepatic inflammation marker, D Hepatic fibrosis marker, and E circulating triglyceride levels in male and female control and Tlcd1/2 double-knockout mice on a Western diet. Petkevicius et al., Nat Commun, 13, 6020 (2022), 10.1038/s41467-022-33735-6
Overall, this study performs research on fatty liver disease and identifies TLCD1/2 proteins as regulators of the fatty acyl composition at the sn-1 position of PE, promoting the incorporation of MUFAs. TLCD1/2 appear to act downstream of MUFA uptake and upstream of LPLAT enzymes, suggesting a potential role in intracellular lipid trafficking.
PE remodelling links Tlcd1/2 to the development of fatty liver disease, with sex-specific protective effects observed in Tlcd1/2 DKO mice. Beyond fatty liver disease, TLCD1 has been associated with hepatocellular carcinoma and poor prognosis, while Tlcd2 influences eosinophil levels during allergic lung inflammation. These findings suggest broader roles for TLCD1/2 in various physiological and disease contexts.
Lipotype lipidomics technology allows for detailed analysis of lipid composition in biological systems, providing insight into how specific lipids such as arachidonic acid and other fatty acids contribute to liver diseases, including MASLD (NAFLD) and MASH (NASH). This supports the discovery of disease biomarkers, therapeutic targets, and lipid-based drug delivery strategies.
Not sure how this works for your case? Let’s talk it through!
AstraZeneca is a global, science-driven biopharmaceutical company focused on advancing innovative treatments that deliver lasting benefits for patients, society, the environment, and its stakeholders.
Newsletter
Get the latest lipidomics articles and publications delivered straight to you.
You are currently viewing a placeholder content from OpenStreetMap. To access the actual content, click the button below. Please note that doing so will share data with third-party providers.

 progression.")

 male mice. The y-axis displays the negative logarithm of p-values from two-tailed multiple t-tests (unadjusted for multiple comparisons).")

 between control and Tlcd1/2 double-knockout male mice (from unpaired two-tailed Student’s t-tests.")
, total liver lipid content, and serum ALT activity in high-fat-diet–fed control and Tlcd1/2 double-knockout male and female mice.")
, B Serum ALT activity, C Hepatic inflammation marker, D Hepatic fibrosis marker, and E circulating triglyceride levels in male and female control and Tlcd1/2 double-knockout mice on a Western diet.")