

A lipid droplet is a very dynamic organelle that plays a key role in managing lipid metabolism, storage, and transport. This specialized organelle can store lipids produced within the cell, helping to maintain organelle and membrane stability, which is crucial for cell survival and resistance to stress. Lipid droplets play an essential role in CNS regeneration and remyelination, and in various diseases like ischemic stroke.
Lipid droplets form at the endoplasmic reticulum (ER) through localized lipogenesis under different conditions such as a nutrient excess. In this way, fatty acids and sterols are stored in lipid droplets in the form of triacylglycerols (TAGs) and cholesteryl esters. This process of TAG buildup happens in the space between endoplasmic reticulum membranes. The accumulated lipids create a lipid lens that expands and pushes the membrane toward the cytosol. Continued growth causes the droplet to bud off into the cytosol. Smaller lipid droplets either merge with larger ones or transfer their lipids to them. Larger lipid droplets remain in the cytosol. When lipid remobilization is triggered, the droplets break down. For example, during starvation, lipid droplets break down triacylglycerols (TAGs) into free fatty acids and glycerol, which are then used for energy production through mitochondrial fatty acids oxidation. In cases when cell membranes need to be produced, products of TAG degradation supply lipids for phospholipid synthesis and maintenance.
 through localized lipogenesis. This process leads to the buildup of triacylglycerols (TAGs) in the space between ER membranes. The accumulated lipids create a lipid lens that expands and pushes the membrane towards the cytosol. Continued growth causes the droplet to bud off into the cytosol. Smaller lipid droplets either merge with larger ones or transfer their lipids to them. Larger lipid droplets remain in the cytosol. When lipid remobilization is triggered, the droplets break down. Fatty acids released from storage lipids can be re-esterified to re-enter the lipid droplet cycle, used in β-oxidation for producing energy, or participate in membrane or hormone synthesis, or other reactions.")
Certain fatty acids can compromise membrane integrity and cell metabolism by altering its permeability through lipotoxicity and oxidative stress. Cells prevent this by transferring these fatty acids into lipid droplets covered by phospholipid monolayer to protect cells. In this monolayer, the hydrophobic tails of phospholipids face inward toward the hydrophobic core of the lipid droplet. In addition to fatty acids incorporating into TAGs in lipid droplets, other lipids, like squalene and zeaxanthin, can migrate into lipid droplets. There is still much to learn about how these lipids are partitioned within lipid droplets.
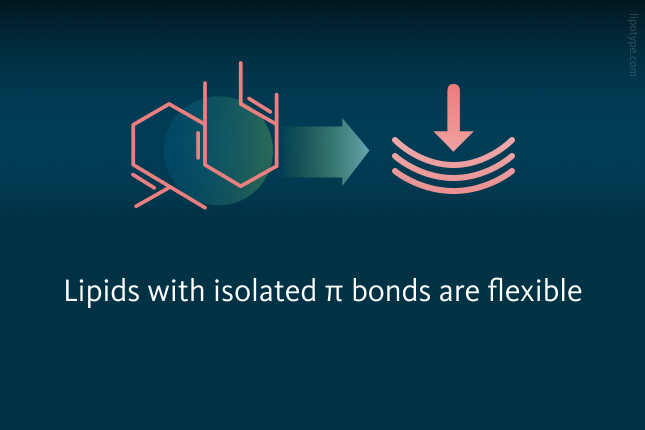
The authors, a group of researchers led by scientists from the Korea Research Institute of Chemical Technology, applied molecular dynamics simulations alongside microbial lipid engineering to evaluate if the structural flexibility of lipids plays a role in their migration into lipid droplets. The π bonds in these lipids may be conjugated or non-conjugated, and this may play a role in their selective diffusion into lipid droplets as well as subsequent partitioning into cytosolic lipid droplets. The scientists started by investigating how the flexibility of lipids influences the selective migration of these lipids into lipid droplets.
 is structurally flexible due to its isolated π bonds. In contrast, zeaxanthin (green) has conjugated π bonds, making it rigid and resulting in minimal conformational changes.")
The chemical structure of the non-saponifiable lipids used in the computational and experimental analyses revealed that squalene is structurally flexible due to its isolated π bonds. In contrast, zeaxanthin has conjugated π bonds, making it rigid and resulting in minimal conformational changes.
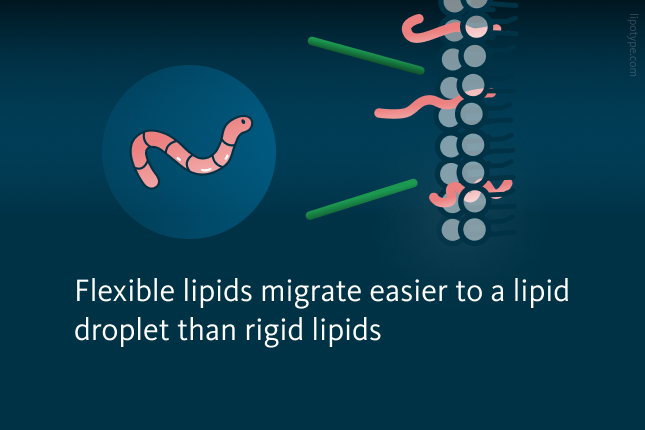
Flexible lipids can easily deform, which lowers the free energy barrier and allows them to pass through TAGs that are tightly packed, affecting lipid droplet composition. In contrast, rigid lipids must significantly rearrange the intertwined TAG chains to penetrate deeply into a lipid droplet. Due to the high free energy barrier, these rigid lipids become stuck in the membrane and cannot access the lipid droplet core.
 boundaries. More flexible lipids easily pass the membrane and migrate to a lipid droplet core.")
By systematically regulating lipid droplet synthesis and degradation pathways in yeast cells, the researchers managed to control the size and number of intracellular lipid droplets. Specifically, the strategy for engineering lipid droplet size and number concentrated on ten different genes associated with the TAG metabolism and membrane transformation.
 of several lipid metabolism related genes and the measured impact on lipid droplet size and number.")
Genes associated with lipid droplet metabolism. A tabular overview for the experimental genetic engineering (overexpression of deletion) of several lipid metabolism related genes and the measured impact on lipid droplet size and number.
Son et al., Nat Commun 13, 3612 (2022), 10.1038/s41467-022-31400-6
To determine if lipid structural flexibility significantly affects their migration and storage in lipid droplets in vivo, the authors investigated the storage patterns of intracellularly produced squalene and zeaxanthin. They found that squalene storage increased as lipid droplet size grew. Notably, squalene storage increased exponentially with the total lipid droplet volume. It seems that lipid droplet metabolism and composition are affected in the following way: as the bulky TAGs are diluted by single-chain squalene, the TAG-packaging LDs may become leaky, promoting further squalene migration and allowing a substantial accumulation of squalene in the more relaxed LDs.
In contrast, the storage capacity for rigid, non-flexible lipids like zeaxanthin and β-carotene did not significantly increase in the engineered yeast cells, even though the net surface area of LDs was increased. These rigid, non-foldable lipids could not be stored within the intracellular LDs and were instead harboured by various membranes, including organelles and plasma membranes.

Lipid profiles of lipid droplets in various yeast cells. Lipid profiles of the engineered yeast cells revealed that total lipid and lipid droplet lipid levels in cells with increased lipid droplet size and cells with increased lipid droplet number were slightly elevated compared to control cells. Lipid droplet lipid composition in these cells and total lipid levels were similar. The relative enrichment of lipids in lipid droplets in these engineered cells is illustrated in mol %. DAG, diacylglycerol; TAG, triacylglycerol; SE, sterol ester; PA, phosphatidic acid; PC, phosphatidylcholine; PE, phosphatidylethanolamine; PG, phosphatidylglycerol; PI, phosphatidylinositol; PS, phosphatidylserine; LPs, lysophospholipids; DCW, dry cell weight.
Son et al., Nat Commun 13, 3612 (2022), 10.1038/s41467-022-31400-6
Rigid, non-flexible zeaxanthin displayed a completely different storage-dependence pattern in the engineered yeast cells compared to flexible squalene. Cells with numerous small lipid droplets stored more zeaxanthin than those with fewer, larger lipid droplets. This suggests that the overall surface area of the lipid droplets is a critical factor for zeaxanthin storage. The authors concluded that the amount of stored zeaxanthin correlated to the surface area of each lipid droplet. Additionally, it is likely that membrane-bound organelles and cell membranes also acted as storage sites for zeaxanthin.
Overall, this study performed the characterization of lipid classes in lipid droplets from various yeast cells. The researchers also concluded that the flexibility of lipids is a key determinant of a lipid’s ability to migrate into lipid droplets.

Lipotype Lipidomics technology offers support in yeast samples and other studies by providing comprehensive lipidomics analyses. It enables detailed profiling of lipid species, including squalene and oxidized squalene, within lipid droplets and other cellular compartments, elucidating their structural characteristics, flexibility, and storage dynamics.
Do you have any questions?
We can answer them!

Lipotype products are provided for Research Use Only. They are not intended for clinical diagnostic purposes and must not be used to inform medical treatment decisions. The content of this article is for scientific and educational purposes only and should not be considered medical advice.



